Electron transfer in proteins
Electron transfer (ET) by proteins is fundamental to all life. Although they essentially operate as single-molecular machines, most of our understanding of these proteins is based upon measurements of bulk material.
But in recent years, single molecule studies generally (including those on proteins) have been made possible by scanning probe-based studies: for example, individual protein molecule unfolding events can be measured with AFM (atomic force microscopy), and single molecule electrical conduction (probing temporal and spatial variations hidden in the bulk) can be studied using STM (scanning tunnelling microscopy) methods. Variations in individual molecule/environment interactions and their dynamics can thus be revealed.
Activities
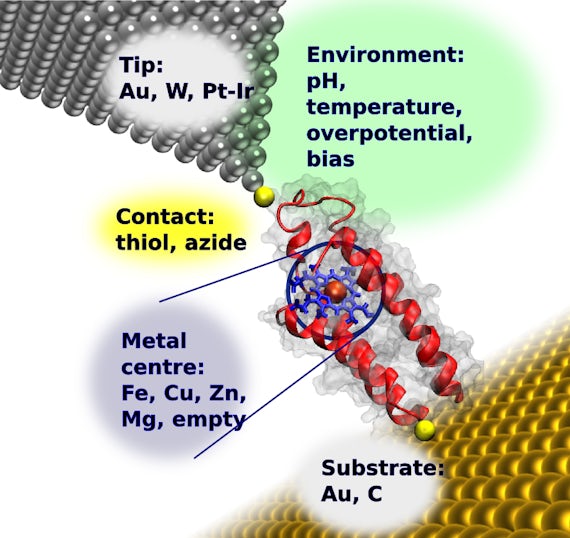
We engineer the redox-active protein cytochrome b562 to introduce pairs of thiol groups at specific places on opposite ends of the molecule. The protein maintains its native (unfolded) form under the experimental conditions. This degree of control of orientation and electrical contacts is a prerequisite for integration into electronic circuits.
Stable protein-gold STM tip electrical contact is made, and current-voltage and current-distance measurements taken.
Proteins with thiol contacts positioned across the protein's short axis have a conductance of (3.48 ± 0.05) × 10-5 G0.
But proteins with thiol groups placed along the long axis reproducibly yields two distinct values of (1.95 ± 0.03) × 10-5 G0 and (3.57 ± 0.11) × 10-5 G0. This indicates that the placement of the asymmetrically located haem within the protein controls electron transfer.
In contrast, the unengineered wild-type cytochrome b562 had conductance values at least 1 order of magnitude less.
Electrochemical gating, i.e. by measuring within a solution and applying a potential to the solution via a third electrode, shows that the conductance can be modulated or "gated". This transistor action originates in the shift of the heme energy levels with potential.
Publications
- Della Pia, E. A. et al., 2012. Direct binding of a redox protein for single-molecule electron transfer measurements. Small 8 (15), pp.2341-2344. (10.1002/smll.201102416)
- Della Pia, E. A. et al. 2012. Redox tuning of cytochrome b562 through facile metal porphyrin substitution. Chemical Communications 48 (86), pp.10624-10626. (10.1039/c2cc34302a)
- Della Pia, E. A. et al. 2012. Fast electron transfer through a single molecule natively structured redox protein. Nanoscale 4 (22), pp.7106-7113. (10.1039/c2nr32131a)
- Della Pia, E. A. et al. 2011. Single-molecule mapping of long-range electron transport for a cytochrome b562 variant. Nano Letters 11 (1), pp.176-182. (10.1021/nl103334q)