
Professor Helen White-Cooper
Director of Research (Innovation and Environment)
School of Biosciences
- White-CooperH@cardiff.ac.uk
- +44 29208 75492
- Sir Martin Evans Building, Room W3.21, Museum Avenue, Cardiff, CF10 3AX
- Available for postgraduate supervision
Overview
Research overview
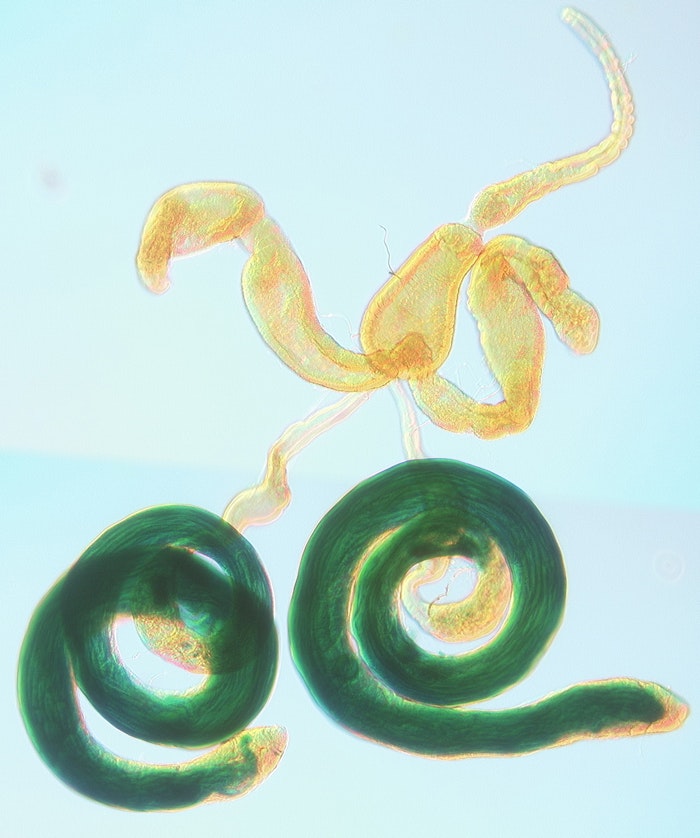
Developmental Genetics - Regulation of Gene Expression in Drosophila Spermatogenesis
Cell differentiation is driven by co-ordinated changes in the gene expression profile of the cell: some genes are switched on, others are switched off. The mature sperm is a highly specialised cell (nearly 2mm long), whose formation from a simple primary spermatocyte involves meiosis to form round spermatids, followed by complex changes in cell architecture to form the final elongated motile sperm. During spermatogenesis there is a dramatic switch in the gene expression profile of male germ-line cells: as they enter the primary spermatocyte stage they activate transcription of a large set of genes required for sperm production. We have identified a set of proteins, encoded by the meiotic arrest genes, that work together to activate this transcriptional programme, and are investigating the composition, activity and evolution of this complex. We recently discovered that another small set of genes is transcribed after meiosis, and that these late transcribed mRNAs localise to a discrete region of the cell. We are studying their transcriptional control, and the mRNA localisation mechanism. Continued production of sperm is maintained via a stem cell system, and we are characterising a transcription factor required for stem cell maintenance.
Publication
2024
- McNamara, C. J., Ant, T. H., Harvey-Samuel, T., White-Cooper, H., Martinez, J., Alphey, L. and Sinkins, S. P. 2024. Transgenic expression of cif genes from Wolbachia strain w AlbB recapitulates cytoplasmic incompatibility in Aedes aegypti. Nature Communications 15, article number: 869. (10.1038/s41467-024-45238-7)
2023
- Raz, A. A. et al. 2023. Emergent dynamics of adult stem cell lineages from single nucleus and single cell RNA-Seq of Drosophila testes. eLife 12, article number: e82201. (10.7554/elife.82201)
2022
- Li, H. et al. 2022. Fly cell atlas: a single-nucleus transcriptomic atlas of the adult fruit fly. Science 375(6584), article number: 2432. (10.1126/science.abk2432)
2021
- van der Graaf, K., Jindrich, K., Mitchell, R. and White-Cooper, H. 2021. Roles for RNA export factor, Nxt1, in ensuring muscle integrity and normal RNA expression in Drosophila.. G3 11(1), article number: jkaa046. (10.1093/g3journal/jkaa046)
2018
- Laktionov, P. P. et al. 2018. Genome-wide analysis of gene regulation mechanisms during Drosophila spermatogenesis. Epigenetics and Chromatin 11, article number: 14. (10.1186/s13072-018-0183-3)
2016
- Sutton, E., Yu, Y., Shimeld, S., White-Cooper, H. and Alphey, L. 2016. Identification of genes for engineering the male germline of Aedes aegypti and Ceratitis capitata. BMC Genomics 17, article number: 948. (10.1186/s12864-016-3280-3)
2014
- Lowe, N. et al. 2014. Analysis of the expression patterns, subcellular localisations and interaction partners of Drosophila proteins using a pigP protein trap library. Development 141(20), pp. 3994-4005. (10.1242/dev.111054)
- Lakitionov, P. P., White-Cooper, H., Maksimov, D. A. and Belyakin, S. N. 2014. Transcription factor Comr acts as a direct activator in the genetic program controlling spermatogenesis in D. melanogaster. Molecular Biology 48(1), pp. 130-140. (10.1134/S0026893314010087)
2013
- Caporilli, S., Yu, Y., Jiang, J. and White-Cooper, H. 2013. The RNA export factor, Nxt1, is required for tissue specific transcriptional regulation. PLOS Genetics 9(6), article number: e1003526. (10.1371/journal.pgen.1003526)
- White-Cooper, H. and Caporilli, S. 2013. Transcriptional and post-transcriptional regulation of Drosophila Germline stem cells and their differentiating progeny. In: Hime, G. and Abud, H. eds. Transcriptional and Translational Regulation of Stem Cells., Vol. 1. Advances in Experimental Medicine and Biology Vol. 786. Dordrecht: Springer, pp. 47-61., (10.1007/978-94-007-6621-1_4)
2012
- White-Cooper, H. 2012. Tissue, cell type and stage-specific ectopic gene expression and RNAi induction in the Drosophila testis. Spermatogenesis 2(1), pp. 11-22. (10.4161/spmg.19088)
2011
- Doggett, K., Jiang, J., Aleti, G. and White-Cooper, H. 2011. Wake-up-call, a lin-52 paralogue, and always early, a lin-9 homologue physically interact, but have opposing functions in regulating testis-specific gene expression. Developmental Biology 355(2), pp. 381-393. (10.1016/j.ydbio.2011.04.030)
- White-Cooper, H. and Davidson, I. 2011. Unique aspects of transcription regulation in male germ cells. Cold Spring Harbor Perspectives in Biology 3(7), article number: a002626. (10.1101/cshperspect.a002626)
2010
- Miles, A., Zhao, J., Klyne, G., White-Cooper, H. and Shotton, D. 2010. OpenFlyData: An exemplar data web integrating gene expression data on the fruit fly Drosophila melanogaster. Journal of Biomedical Informatics 43(5), pp. 752-761. (10.1016/j.jbi.2010.04.004)
- White-Cooper, H. and Bausek, N. 2010. Evolution and spermatogenesis. Philosophical Transactions of the Royal Society of London Series B Biological Sciences 365(1546), pp. 1465-1480. (10.1098/rstb.2009.0323)
- Fu, G. et al. 2010. Female-specific flightless phenotype for mosquito control. Proceedings of the National Academy of Sciences of the United States of America 107(10), pp. 4550-4554. (10.1073/pnas.1000251107)
- Zhao, J., Klyne, G., Benson, E., Gudmannsdottir, E., White-Cooper, H. and Shotton, D. 2010. FlyTED: The Drosophila testis gene expression database. Nucleic Acids Research 38(S1), pp. D710-D715. (10.1093/nar/gkp1006)
- White-Cooper, H. 2010. Molecular mechanisms of gene regulation during Drosophila spermatogenesis. Reproduction 139(1), pp. 11-21. (10.1530/REP-09-0083)
2009
- Morris, C., Benson, E. and White-Cooper, H. 2009. Determination of gene expression patterns using in situ hybridization to Drosophila testes. Nature Protocols 4(12), pp. 1807-1819. (10.1038/nprot.2009.192)
- Sato, A., White-Cooper, H., Doggett, K. and Holland, P. W. H. 2009. Degenerate evolution of the hedgehog gene in a hemichordate lineage. Proceedings of the National Academy of Sciences of the United States of America 106(18), pp. 7491-7494. (10.1073/pnas.0810430106)
- White-Cooper, H. 2009. Studying how flies make sperm-Investigating gene function in Drosophila testes. Molecular and Cellular Endocrinology 306(1-2), pp. 66-74. (10.1016/j.mce.2008.11.026)
2008
- Barreau, C., Benson, E., Gudmannsdottir, E., Newton, F. and White-Cooper, H. 2008. Post-meiotic transcription in Drosophila spermatogenesis. Development 135(11), pp. 1897-1902. (10.1242/dev.021949)
- Barreau, C., Benson, E. and White-Cooper, H. 2008. Comet and cup genes in Drosophila spermatogenesis: the first demonstration of post-meiotic transcription. Biochemical Society Transactions 36(3), pp. 540-542. (10.1042/BST0360540)
- Kirchner, J., Vissi, E., Gross, S., Szoor, B., Rudenko, A., Alphey, L. and White-Cooper, H. 2008. Drosophila Uri, a PP1alpha binding protein, is essential for viability, maintenance of DNA integrity and normal transcriptional activity. BMC Molecular Biology 9: 36 (10.1186/1471-2199-9-36)
- Townley, H. E., Parker, A. R. and White-Cooper, H. 2008. Exploitation of diatom frustules for nanotechnology: tethering active biomolecules. Advanced Functional Materials 18(2), pp. 369-374. (10.1002/adfm.200700609)
- Nair-Roberts, R. G., Chatelain-Badie, S. D., Benson, E., White-Cooper, H., Bolam, J. P. and Ungless, M. A. 2008. Stereological estimates of dopaminergic, GABAergic and glutamatergic neurons in the ventral tegmental area, substantia nigra and retrorubral field in the rat. Neuroscience 152(4), pp. 1024-31. (10.1016/j.neuroscience.2008.01.046)
2007
- Jiang, J., Benson, E., Bausek, N., Doggett, K. and White-Cooper, H. 2007. Tombola, a tesmin/TSO1 family protein, regulates transcriptional activation in the Drosophila male germline and physically interacts with Always early. Development 134, pp. 1549-1559. (10.1242/dev.000521)
2004
- L, P., J, J., B, B., M, H., E, B., MT, F. and White-Cooper, H. 2004. Regulation of transcription of meiotic cell cycle and terminal differentiation genes by the testis-specific Zn finger protein matotopetli. Development 131, pp. 1691-1702. (10.1242/dev.01032)
- Korenjak, M. et al. 2004. Native E2F/RBF complexes contain Myb-interacting proteins and repress transcription of developmentally controlled E2F target genes. Cell 19(2), pp. 181-193. (10.1016/j.cell.2004.09.034)
2003
- Jiang, J. and White-Cooper, H. 2003. Transcriptional activation in Drosophila spermatogenesis involves the mutually dependent function of aly and a novel meiotic arrest gene cookie monster. Development 130(3), pp. 563-573. (10.1242/dev.00246)
Articles
- McNamara, C. J., Ant, T. H., Harvey-Samuel, T., White-Cooper, H., Martinez, J., Alphey, L. and Sinkins, S. P. 2024. Transgenic expression of cif genes from Wolbachia strain w AlbB recapitulates cytoplasmic incompatibility in Aedes aegypti. Nature Communications 15, article number: 869. (10.1038/s41467-024-45238-7)
- Raz, A. A. et al. 2023. Emergent dynamics of adult stem cell lineages from single nucleus and single cell RNA-Seq of Drosophila testes. eLife 12, article number: e82201. (10.7554/elife.82201)
- Li, H. et al. 2022. Fly cell atlas: a single-nucleus transcriptomic atlas of the adult fruit fly. Science 375(6584), article number: 2432. (10.1126/science.abk2432)
- van der Graaf, K., Jindrich, K., Mitchell, R. and White-Cooper, H. 2021. Roles for RNA export factor, Nxt1, in ensuring muscle integrity and normal RNA expression in Drosophila.. G3 11(1), article number: jkaa046. (10.1093/g3journal/jkaa046)
- Laktionov, P. P. et al. 2018. Genome-wide analysis of gene regulation mechanisms during Drosophila spermatogenesis. Epigenetics and Chromatin 11, article number: 14. (10.1186/s13072-018-0183-3)
- Sutton, E., Yu, Y., Shimeld, S., White-Cooper, H. and Alphey, L. 2016. Identification of genes for engineering the male germline of Aedes aegypti and Ceratitis capitata. BMC Genomics 17, article number: 948. (10.1186/s12864-016-3280-3)
- Lowe, N. et al. 2014. Analysis of the expression patterns, subcellular localisations and interaction partners of Drosophila proteins using a pigP protein trap library. Development 141(20), pp. 3994-4005. (10.1242/dev.111054)
- Lakitionov, P. P., White-Cooper, H., Maksimov, D. A. and Belyakin, S. N. 2014. Transcription factor Comr acts as a direct activator in the genetic program controlling spermatogenesis in D. melanogaster. Molecular Biology 48(1), pp. 130-140. (10.1134/S0026893314010087)
- Caporilli, S., Yu, Y., Jiang, J. and White-Cooper, H. 2013. The RNA export factor, Nxt1, is required for tissue specific transcriptional regulation. PLOS Genetics 9(6), article number: e1003526. (10.1371/journal.pgen.1003526)
- White-Cooper, H. 2012. Tissue, cell type and stage-specific ectopic gene expression and RNAi induction in the Drosophila testis. Spermatogenesis 2(1), pp. 11-22. (10.4161/spmg.19088)
- Doggett, K., Jiang, J., Aleti, G. and White-Cooper, H. 2011. Wake-up-call, a lin-52 paralogue, and always early, a lin-9 homologue physically interact, but have opposing functions in regulating testis-specific gene expression. Developmental Biology 355(2), pp. 381-393. (10.1016/j.ydbio.2011.04.030)
- White-Cooper, H. and Davidson, I. 2011. Unique aspects of transcription regulation in male germ cells. Cold Spring Harbor Perspectives in Biology 3(7), article number: a002626. (10.1101/cshperspect.a002626)
- Miles, A., Zhao, J., Klyne, G., White-Cooper, H. and Shotton, D. 2010. OpenFlyData: An exemplar data web integrating gene expression data on the fruit fly Drosophila melanogaster. Journal of Biomedical Informatics 43(5), pp. 752-761. (10.1016/j.jbi.2010.04.004)
- White-Cooper, H. and Bausek, N. 2010. Evolution and spermatogenesis. Philosophical Transactions of the Royal Society of London Series B Biological Sciences 365(1546), pp. 1465-1480. (10.1098/rstb.2009.0323)
- Fu, G. et al. 2010. Female-specific flightless phenotype for mosquito control. Proceedings of the National Academy of Sciences of the United States of America 107(10), pp. 4550-4554. (10.1073/pnas.1000251107)
- Zhao, J., Klyne, G., Benson, E., Gudmannsdottir, E., White-Cooper, H. and Shotton, D. 2010. FlyTED: The Drosophila testis gene expression database. Nucleic Acids Research 38(S1), pp. D710-D715. (10.1093/nar/gkp1006)
- White-Cooper, H. 2010. Molecular mechanisms of gene regulation during Drosophila spermatogenesis. Reproduction 139(1), pp. 11-21. (10.1530/REP-09-0083)
- Morris, C., Benson, E. and White-Cooper, H. 2009. Determination of gene expression patterns using in situ hybridization to Drosophila testes. Nature Protocols 4(12), pp. 1807-1819. (10.1038/nprot.2009.192)
- Sato, A., White-Cooper, H., Doggett, K. and Holland, P. W. H. 2009. Degenerate evolution of the hedgehog gene in a hemichordate lineage. Proceedings of the National Academy of Sciences of the United States of America 106(18), pp. 7491-7494. (10.1073/pnas.0810430106)
- White-Cooper, H. 2009. Studying how flies make sperm-Investigating gene function in Drosophila testes. Molecular and Cellular Endocrinology 306(1-2), pp. 66-74. (10.1016/j.mce.2008.11.026)
- Barreau, C., Benson, E., Gudmannsdottir, E., Newton, F. and White-Cooper, H. 2008. Post-meiotic transcription in Drosophila spermatogenesis. Development 135(11), pp. 1897-1902. (10.1242/dev.021949)
- Barreau, C., Benson, E. and White-Cooper, H. 2008. Comet and cup genes in Drosophila spermatogenesis: the first demonstration of post-meiotic transcription. Biochemical Society Transactions 36(3), pp. 540-542. (10.1042/BST0360540)
- Kirchner, J., Vissi, E., Gross, S., Szoor, B., Rudenko, A., Alphey, L. and White-Cooper, H. 2008. Drosophila Uri, a PP1alpha binding protein, is essential for viability, maintenance of DNA integrity and normal transcriptional activity. BMC Molecular Biology 9: 36 (10.1186/1471-2199-9-36)
- Townley, H. E., Parker, A. R. and White-Cooper, H. 2008. Exploitation of diatom frustules for nanotechnology: tethering active biomolecules. Advanced Functional Materials 18(2), pp. 369-374. (10.1002/adfm.200700609)
- Nair-Roberts, R. G., Chatelain-Badie, S. D., Benson, E., White-Cooper, H., Bolam, J. P. and Ungless, M. A. 2008. Stereological estimates of dopaminergic, GABAergic and glutamatergic neurons in the ventral tegmental area, substantia nigra and retrorubral field in the rat. Neuroscience 152(4), pp. 1024-31. (10.1016/j.neuroscience.2008.01.046)
- Jiang, J., Benson, E., Bausek, N., Doggett, K. and White-Cooper, H. 2007. Tombola, a tesmin/TSO1 family protein, regulates transcriptional activation in the Drosophila male germline and physically interacts with Always early. Development 134, pp. 1549-1559. (10.1242/dev.000521)
- L, P., J, J., B, B., M, H., E, B., MT, F. and White-Cooper, H. 2004. Regulation of transcription of meiotic cell cycle and terminal differentiation genes by the testis-specific Zn finger protein matotopetli. Development 131, pp. 1691-1702. (10.1242/dev.01032)
- Korenjak, M. et al. 2004. Native E2F/RBF complexes contain Myb-interacting proteins and repress transcription of developmentally controlled E2F target genes. Cell 19(2), pp. 181-193. (10.1016/j.cell.2004.09.034)
- Jiang, J. and White-Cooper, H. 2003. Transcriptional activation in Drosophila spermatogenesis involves the mutually dependent function of aly and a novel meiotic arrest gene cookie monster. Development 130(3), pp. 563-573. (10.1242/dev.00246)
Book sections
- White-Cooper, H. and Caporilli, S. 2013. Transcriptional and post-transcriptional regulation of Drosophila Germline stem cells and their differentiating progeny. In: Hime, G. and Abud, H. eds. Transcriptional and Translational Regulation of Stem Cells., Vol. 1. Advances in Experimental Medicine and Biology Vol. 786. Dordrecht: Springer, pp. 47-61., (10.1007/978-94-007-6621-1_4)
Research
Regulation of Gene Expression in Drosophila Spermatogenesis
Cell differentiation is driven by co-ordinated changes in the gene expression profile of the cell: some genes are switched on, others are switched off. One of the most astonishing developmentally regulated changes in cell morphology occurs in spermatogenesis. The mature sperm is a highly specialised cell, whose formation from a simple primary spermatocyte involves an unusual cell division (meiosis) to form a round spermatid, followed by complex changes in the cell architecture to form the final elongated motile sperm. These differentiation events require many gene products used at no other time in development. Underlying this there is a dramatic switch in the gene expression profile of male germ-line cells: as they enter the primary spermatocyte stage they activate transcription of a large set of genes required for sperm production. We have recently discovered that another small set of genes is transcribed after meiosis, in elongating spermatids, and are investigating their role in sperm function.
The meiotic arrest loci
The meiotic arrest class of Drosophila genes regulate transcription in spermatogenesis; specifically they are required for activation of expression of numerous genes required for spermatid differentiation. A series of micro-array experiments revealed that approximately half of all Drosophila protein coding genes are expressed in testes, and that 15-20% of these are regulated by our genes, ie, the meiotic arrest genes control transcription of up to 10% of all Drosophila protein coding genes. We cloned and characterised five of these meiotic arrest genes (aly, comr, achi/vis, topi and tomb). aly and comr have unknown functions, topi, tomb and achi/vis encode DNA binding proteins. In normal primary spermatocytes all the meiotic arrest proteins are chromatin associated, consistent with their role in gene-expression regulation. Their localisation in different mutant backgrounds varies: e.g. aly function is required for the nuclear localisation of Comr, and vice versa, indicating that formation and localisation of an active complex is highly regulated. We are currently investigating the formation of the complex, and its activity at target promoters.
Gene function in spermatogenesis
In 1998 I drew a model of the function of the meiotic arrest genes, in which I postulated that aly would regulate many genes required for spermatid differentiation. This model was based on expression data for about 20 genes. The array experiment was designed to test the model with many more genes. However, the main observation revealed by the array analysis is that we know very little about gene function in Drosophila spermatogenesis. I could initially not answer my question "do the meiotic arrest loci predominantly regulate genes required for spermiogenesis, and not genes required before meiosis?" because the function of the majority of genes that changed significantly in the mutants was not known. We therefore began a large-scale functional-genetics project in which we are using RNA in situ hybridisation to describe the expression pattern, and genetic regulation, of more than 1000 genes in testes. We have examined the expression patterns of about 1200 genes, and it seems my initial prediction was correct – genes that act after meiosis tend to be regulated by the meiotic arrest genes, whereas genes that act earlier tend not to require the meiotic arrest genes for their expression.
Post-meiotic gene expression in Drosophila
It has been generally accepted that there is no post-meiotic transcription in Drosophila spermatogenesis. However, in our in situ hybridisation project, we have identified about 25 genes that are transcribed during the elongation stage of spermatid development. Moreover, their transcripts are localised to the distal ends of the spermatids. The localised transcripts fall into two classes - "cups" and "comets". A mutation in one of the comet genes, scotti, is male sterile, with defects late in spermatogenesis. We are using in vivo reporter constructs and genetic analysis to examine the mechanisms controlling the mRNA and protein localisations and functions.
Watch a video about why sperm are so interesting
Funding
Research in my lab is funded by BBSRC.
Group members
- Dr Saurabh Chaudhary
- Mrs Sabrina Williams
- Dr Fiona Messer
Postgraduate research students
- Miss Dana Jackson
- Miss Cristina Fernadez Garcia
Teaching
I teach fundamentals of genetics and mutations in the year 1 "Genetics and Evolution" module. This includes lectures and a lab practical session on Drosophila genetics.
I teach more on mutations as well as recombination and transgenic animal technologies in "Molecular Biology of the Gene" in year 2. This includes lectures and a lab practical session on DNA damage and repair.
In yr3 "Current Topics in Development, Stem cells and Repair" I lecture on the importance of chromatin and transcriptional regulation in developmental biology, particularly in the context of germ cell specification and function.
I welcome final year project students in the group - typically supervising 4-6 BSc students on lab or data analysis projects, and one Integrated Master's final dissertation student.
Biography
I graduated from Cambridge University with a BA in Natural Sciences (Zoology) in 1990, then went to Dundee University to carry out PhD research on the regulation of cell division in fruit flies. In 1995 I went to Stanford University in California for a period of post-doctoral research, concentrating on the role of specific genes co-ordination of various cellular events during sperm production in flies. In 1998 I moved to Oxford to set up my own lab, initially as a departmental lecturer, and in 2001 earned Royal Society University Research Fellowship. I continued to focus on spermatogenesis in Drosophila, specifically looking at regulation and function of testis specific genes. In April 2008 I moved to Cardiff University, to take up a position as a Senior Lecturer, continuing with the fly testis research. I was promoted to Reader in 2011 and to Professor in 2014.
Committees and reviewing
- Chaired the BBSRC BBR grant committee in 2021 and 2022
- Member of the BBSRC sLOLA Outline stage assessment panel in 2021
- Grant reviewer for BBSRC responsive mode
- Have reviewed papers for numerous journals, including for PLOS genetics and G3
Supervisions
The outlines of two potential projects available now are listed below. If you are interested in studying for a higher degree in my lab we can also work together to generate a project outline that is of mutual interest.
Transcriptional regulation in Drosophila melanogaster testes
The process of sperm production in animals requires activation of many genes that are not used in any other tissues. Indeed, more than 10% of the protein coding genes in both the human and fly genomes are expressed exclusively in the testis. In our lab we investigate the transcriptional controls that generate this testis specificity – both keeping the genes off in other tissues and (the main focus of the lab) activating them in spermatocytes in testes.
A critical, conserved, transcriptional regulatory complex activates expression of many genes in spermatocytes in Drosophila melanogaster. We have mutant alleles of most of the components and have detailed phenotypic analysis, including for example RNAseq to identify transcriptional changes. We have additionally identified a putative novel component of the complex, via protein-interaction assays with known complex subunits. The protein localizes to chromatin in the male germline, consistent with a role in gene expression. We are using CRISPR mutagenesis to generate knock out alleles of this gene. This project will involve analysis of the mutants, initially aiming to determine how the phenotype compares to known pathway components. Further analysis will reveal how the protein interacts with, and modulates the function of, the core complex. The gene has recently evolved, and a cross species comparison will reveal how new components can integrate into established regulatory modules.
Likely Techniques
- Molecular biology – DNA and RNA extraction, PCR, RT-PCR, Q-RT-PCR. Plasmid construction.
- DNA-protein interaction analysis – ChIP seq.
- RNA seq – sample preparation, library construction, bioinformatics.
- Cytology and microscopy of testes; immunostaining; RNA in situ hybridisation.
- Drosophila genetics, including CRISPR tagging, transgenesis and inducible expression systems.
Transcriptional control of sperm heteromorphy:
Normal sperm production and function is critical for reproduction and thus species survival. An important aspect of reproduction is the performance of sperm once they enter the female reproductive tract - they are exposed to a hostile environment (females make spermicides), and have to compete with sperm from other males to achieve fertilisation of the egg. In many species this has resulted in the males making two or more different types of sperm. One type is capable of fertilisation, while the other(s) act as helpers, by counteracting the spermicide and by dislodging sperm from other males. This is an important evolutionary adaptation, but leaves us as molecular biologists wondering "how do the males manage to make these different sperm types?".
Drosophila pseudoobscura males make three types of sperm (two short, one long), of which just one (long) is capable of fertilising eggs. We have identified a set of genes that are expressed more highly in precursors of either long or short sperm, as well as some regulators that probably activate and/or repress these genes in the appropriate cells.
We are exploiting the CRISPR-CAS9 system for genome editing in D. pseudoobscura, to insert tags onto proteins of interest and thus investigate protein localisation and function, or to make knock out lines. We are currently testing this system, and also using conventional transgeneis to generate lines expressing tagged proteins. You will investigate the function of these factors – what target sites do they bind in the genome (ChIP-seq with tagged proteins); what is the effect of mis-expressing them in the “wrong” cells (transgenics, single cell analysis); what is the effect of mutating them (CRISPR knock outs followed by RNAseq)?
Likely Techniques
- Drosophila genetics, including injection and generation of transgenics.
- Molecular biology – PCR, Q-PCR, RT-PCR, cloning, CRISPR construct designs and application.
- Cell biology – in situ hybridisation, immunostaining, cytology.
- RNAseq and ChIP-seq, including bioinformatics analysis.
Research themes
Specialisms
- Animal developmental and reproductive biology
- Developmental genetics
- Molecular biology
- Cell Biology